Yoneda R.*, Satoh Y.*, Yoshida I., Kawamura S., Kotani T., and Kimura A.P.
*equally contributed
A genomic region transcribed into a long noncoding RNA interacts with the Prss42/Tessp-2 promoter in spermatocytes during mouse spermatogenesis, and its flanking sequences can function as enhancers
Molecular Reproduction and Development 83: 541-557
PubMed HUSCAP
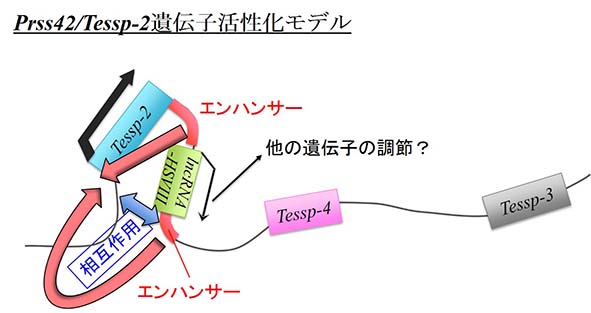
変乆偼儅僂僗惛憙偺尭悢暘楐夁掱偱堦師惛曣嵶朎偵偍偄偰摿堎揑偵揮幨妶惈壔偝傟傞3偮偺僾儘僥傾乕僛堚揱巕丄Prss42/Tessp-2丄 Prss44/Tessp-4丄Prss43/Tessp-3傪摨掕偟偰丄偙傟傜偑尭悢暘楐偺恑峴偲惗怋嵶朎偺惗懚偵廳梫側栶妱傪壥偨偡偙偲傪柧傜偐偵偟 傑偟偨乮Yoneda et al., 2013乯丅崱夞偼偙傟傜偺堚揱巕偑堦師惛曣嵶朎偱摿堎揑偵妶惈壔偝傟傞儊僇僯僘儉傪夝愅偟傑偟偨丅傑偢儅僂僗惛憙偺惗怋嵶朎偲 娞憻偺嵶朎傪廤傔偰DNase I 崅姶庴惈椞堟偲僲儞僐乕僨傿儞僌揮幨傪挷傋傑偟偨丅DNase I 崅姶庴惈椞堟偲偄偆偺偼丄偦偺嵶朎丒慻怐偱僋儘儅僠儞偺娚傫偱偄傞椞堟偺偙偲傪巜偟丄捠忢偼揮幨挷愡偵廳梫側攝楍偲側偭偰偄傑偡丅夝愅偺寢壥丄惗怋嵶朎偺 傒偱DNase I 崅姶庴惈椞堟偲側偭偰偄傞偺偑Prss42/Tessp-2僾儘儌乕僞乕偩偗偱偁傞偙偲偑傢偐傝傑偟偨乮Fig. 1乯丅堦曽偱丄Prss42/Tessp-2偺壓棳偐傜惛憙摿堎揑側僲儞僐乕僨傿儞僌揮幨偑偁傞偙偲偑傢偐傝丄偙偺揮幨嶻暔傪僋儘乕僯儞僌偟偰 lncRNA-HSVIII偲柤偯偗傑偟偨乮Fig. 2乯丅in situ hybridization夝愅偺寢壥丄lncRNA-HSVIII偼僷僉僥儞婜偲屇偽傟傞帪婜偵偼堦師惛曣嵶朎偺妀偵嬊嵼偟偰偄傑 偟偨偑丄偦偺屻嵶朎幙傊偲嬊嵼傪曄壔偝偣偰惛嵶朎傑偱敪尰偟偰偄傞偙偲偑傢偐傝傑偟偨乮Fig. 3乯丅変乆偼偙偺lncRNA-HSVIII偑3偮偺Prss/Tessp堚揱巕偺偳傟偐傪挷愡偡傞壜擻惈傪峫偊偰丄lncRNA-HSVIII偺揮幨偝 傟傞僎僲儉椞堟偲憡屳嶌梡偡傞椞堟傪挷傋傑偟偨丅Chromosome Confromation Capture朄偵傛偭偰夝愅偟偨偲偙傠丄lncRNA-HSVIII偑揮幨偝傟傞壓棳偺僎僲儉攝楍偑丄惛曣嵶朎偵偍偄偰Prss42/Tessp-2僾 儘儌乕僞乕偲憡屳嶌梡偟偰偄傞偙偲偑傢偐傝傑偟偨乮Fig. 4乯丅偦偙偱丄lncRNA-HSVIII揮幨椞堟偲偦偺忋棳壓棳傪娷傓5.8 kb偺僎僲儉攝楍傪Prss42/Tessp-2僾儘儌乕僞乕偱嬱摦偡傞儖僔僼僃儔乕僛堚揱巕偵偮側偄偩偲偙傠丄偙偺5.8 kb偺攝楍偑Prss42/Tessp-2僾儘儌乕僞乕妶惈傪忋徃偝偣傞偙偲偑傢偐傝傑偟偨乮Fig. 5乯丅偲偙傠偑丄偙偺夝愅偵偍偄偰5.8 kb偺攝楍偐傜lncRNA-HSVIII偑揮幨偝傟偰偄側偄偙偲傕傢偐傝乮Fig. 6乯丄僾儘儌乕僞乕妶惈偺忋徃偑5.8 kb攝楍偵娷傑傟傞僄儞僴儞僒乕妶惈偵傛傞傕偺偱偁傞偙偲偑梊憐偝傟傑偟偨丅偦偙偱丄偙偺5.8 kb攝楍傪3偮偵暘偗偰Prss42/Tessp-2僾儘儌乕僞乕偵懳偡傞僄儞僴儞僒乕妶惈傪挷傋偨偲偙傠丄lncRNA-HSVIII揮幨椞堟偺忋棳壓 棳攝楍偲傕偵僄儞僴儞僒乕妶惈傪帩偮偙偲偑柧傜偐偵側傝傑偟偨乮Fig. 7乯丅偮傑傝丄lncRNA-HSVIII帺懱偼Prss/Tessp堚揱巕偺妶惈壔偵婡擻偟偰偄側偄壜擻惈偑崅偄偐傢傝偵丄偦偺廃曈攝楍偑惛曣嵶朎摿堎 揑側僄儞僴儞僒乕偲偟偰Prss42/tessp-2堚揱巕偺妶惈壔婡擻偟偆傞偙偲偑柧傜偐偵側傝傑偟偨丅惛憙偵偍偗傞尭悢暘楐夁掱偱偼丄摿偵堦師惛曣嵶 朎偱懡偔偺堚揱巕偑堦惸偵妶惈壔偝傟傞偙偲偑抦傜傟偰偄傑偡偑丄偦偺暘巕儊僇僯僘儉偼傛偔傢偐偭偰偍傜偢丄杮尋媶偱偼偦偺儊僇僯僘儉偺堦抂傪柧傜偐偵偟偨 偙偲偵側傝傑偡丅