植物のたくましさ
道ばたの草木を見た時,「たくましい」と感じたことはないでしょうか。当然,植物は「動かない」生き物であり,何らかの災厄から移動して逃れることはできません。草木はそのような災厄(ストレス)をその場で克服しつつ子孫を残しているわけで,私は畏怖を持って植物を見ています。「雑草は切っても切っても生えてくる」ということは一つの植物のたくましさでしょう。これは頂芽優性と呼ばれる現象で,一番先端にある茎頂を失うと,その下で休眠している腋の茎頂が発達を開始し,新しい芽や葉が生えてくるのです。一方,庭仕事をされたことがある方は「雑草を抜いたのに,放置した場所でまだ生えている」ということを経験されたかもしれません。これはもう一つのたくましさであり,根が切断されても枯死する前に植物体が根を急速に再生させ,その場で固着生活を継続させた結果なのです。前述の頂芽優性については,18世紀に進化論で有名なチャールズ・ダーウィンとその息子が最初に報告して以来,長年研究されてきました。近年,頂芽優性を制御しているのはストリゴラクトンという新しい植物ホルモンだということもわかってきました。一方,切断された根がいかに再生してくるのかということについてはあまり研究がありませんでした。というのは土の中で目視できない根の傷害応答はあまり注目されず,根が再生するメカニズムについての研究はほとんどありませんでした。さらに根は自発的に枝分かれしていつも新たな根(側根)を作るので,自発的に作られる側根と傷害によって誘導される側根の区別が難しいことが,研究を困難にしていました。そこで私たちはこの問題に取り組み,傷害による植物根の再生過程に関わる因子が植物ホルモンのオーキシンであること,オーキシン合成の誘導,さらにオーキシンの極性輸送によって植物の根は再生することを明らかにしました。
偶然を見逃さない
生命科学における基礎研究の目標は基本的に「メカニズムの解明」です。解明するためには生物を見れば良いわけですが,見ただけでは解明したということにはなりません。対象とする現象及びそのメカニズムを人に伝達できる「文字」で表したとき,初めて「解明した」ということになります。また,研究目標の設定は最重要項目で,何を・どこまで・いつまでに・解明するのかというプランを立てなければなりません。目標の設定は明確であるべきなのですが,予定した目標に固執するべきではなく目標が明らかである限りは研究の視野は広く持っている必要があります。すべての研究は非常に単純なプロセスに従って実行されます。つまり,ある現象(事実・証拠)を理解し,そのメカニズムの仮説を立て,それを検証する(実験)ということです。生命科学の多くは実験科学で,研究とは実験であるかのような印象がありますが,実際には実験結果をどのように理解し,それを証拠として仮説を構築し,検証するための実験計画を立てているので,実験は時間がかかりますが研究のすべてではありません。綿引研ではこの作業プロセスに例外を作りません。つまり,実験結果を否定(失敗?)せず,結果を理解することに努めています。根切り応答の研究もこのルールに従い,想定した結果とは異なる現象を発見したのです。 想定しなかった結果とは発見が含まれている原石のようなものなのです。
当時大学院生だった徐冬暘(シュドンヤン)さんはある日,別の研究で根を途中で切断したシロイヌナズナを観察していました。そのシロイヌナズナは突然変異体で主根は伸びるけれども側根は出てこないはずだったのですが,あろうことか側根がフサフサと野生型と同じぐらい出ていたのです。彼女はこれを見逃さず,すぐ私に報告してくれました(研究の視野と現象の理解)。使った突然変異体はオーキシン信号伝達経路に異常があるmsg2/iaa19という変異体だったのですが,同様の経路に異常がある変異体で試験しても,本来,側根が出ないはずなのに側根が出てくるものがありました(現象の確認)。そこで慎重に無傷コントロールと根切りした野生型植物を4日後に比較すると,わずかですが確かに側根が増えていることがわかりました。この小さな差が今まで側根が増える現象に気づけなかった原因だろうと考えています。さらに興味深いことに,切断した植物の側根はコントロールより長い,つまり成長が早いということもわかりました。根の総延長を測定するとコントロールと根切りした植物はなんと同じだったのです。地上部と根の量は植物種で一定であるというshoot-root-ratioという現象が古くから知られています。もしかしたら根切りした植物は根の成長を促進させて,shoot-root-ratioを回復させているのかもしれません。側根数が増える現象をRoot-Cutting induced lateral root Number (RCN)と呼び,側根の成長を促進する現象をRoot-Cutting induced lateral root Growth (RCG)と名付けました。今回の論文発表では側根数が増えるRCNについてそのメカニズムを明らかにしました。

無処理のシロイヌナズナと生育途中で主根を切ったシロイヌナズナの様子(左図)及び側根数,側根の成長速度のグラフ(右図)。無処理の方が主根は長くなるが,根切りをした方が側根が多く,成長速度も速くなっていることがわかる。(左図は処理後4日目の写真。赤矢印は主根を切った位置)。
エビデンスから仮説を立て,実験による検証を行う
自発的な側根の形成にはオーキシンが重要な働きを持っていることが詳しく判っているので,私たちは根の再生にオーキシンがどのように関わっているのか?というアプローチを行いました(仮説の検証)。若い植物にオーキシンの極性輸送を阻害する薬(NPAなどの農薬)を投与すると,新しい側根は全くできず主根だけになります。しかし大変驚いたことに,オーキシンの極性輸送を阻害するいろいろな薬を与えても,根を切るとまたもや側根が出てきました。そこでオーキシン信号伝達経路が機能しているのかを遺伝子発現解析で調べてみると,根切りによって確かにオーキシン応答が促進されていました。次に疑ったのはオーキシン合成です。そこでオーキシン合成を阻害する試薬を試したところ,RCNが顕著に抑制されました。さらにオーキシン合成に関する11種類の遺伝子の破壊株を調べたところ,YUCCA9という遺伝子に異常がある変異体はRCNが顕著に抑制されていました。また根切りによってYUCCA9遺伝子の発現量は根切り後,急速に増加し2時間でピークに達している,早い応答であることが判りました。これらから,根切りがYUCCA9遺伝子を誘導することで,根のオーキシン量を増やしていると予想されました。そこで帝京大学理工学部・朝比奈雅志准教授の研究グループと共同研究を行い,根のオーキシン量を測定したところ,根切りによって実際に増加していること,yucca9変異体ではそのような増加は見られないを確認し,YUCCA9遺伝子が根切り応答に必要な遺伝子であるということが明らかになりました。一方,オーキシン合成を阻害する試薬,YUCASINとNPAを同時投与すると側根は全くできないどころか,根切りでも側根の発生は抑制されました。さらにオーキシンの極性輸送に関する変異体にYUCASINを投与すると,野生型より顕著に側根が減少すること,根切りによってオーキシンの極性輸送体遺伝子の発現は上昇することから,根切りによるRCNにはオーキシン合成の促進と同時に極性輸送が必要であると結論付けられました。

本研究で明らかになったメカニズム。根切りをすることでYUCCA9遺伝子が活性化,オーキシンの合成等を経て,側根が作られたり発達したりする。ただし,根切りがどのようなシグナルを引き起こしてYUCCA9を活性化させているかはまだ同定できておらず,今後の研究が待たれる。
根切り応答研究は始まったばかり
植物にとって根の傷害は脱水という形で直ちに地上部に影響してしまう緊急事態であり,根を切られた植物はできるだけ早く根を再生しようとする性質があります。園芸ではこのような植物の性質をうまく利用しています。たとえば世界中で親しまれている日本伝統の園芸芸術,盆栽作りでは慎重に根の剪定(根切り)を行います。根切りされた植物は,水や養分を効率よく吸収できる若い根を限られた空間(鉢)で再生することで,健全かつ小さな植物である盆栽になります。今回の発見は,このような園芸技術としてしばしば利用されている植物根の再生について,その分子メカニズムを解き明かしました。根切りについては農学的見地から効率的な方法が研究されてきましたが,そのメカニズムが明らかになったことで様々な応用が期待されます。また,本研究室ではRCGのメカニズムについても研究が進行中であり,成果を発表する予定です。最大の謎は根切りによって何がYUCCA9遺伝子を誘導するのか,RCNはRCGと同じものなのかという点であり,このアプローチも進行中です。

このような盆栽は園芸家の努力と植物がもつ「たくましさ」の共同作業によってのみ完成する。
Dongyang Xu, Jiahang Miao, Emi Yumoto, Takao Yokota, Masashi Asahina, Masaaki Watahiki “YUCCA9-mediated Auxin Biosynthesis and Polar Auxin Transport Synergistically Regulate Regeneration of Root Systems Following Root Cutting”, Plant and Cell Physiology,(2017) https://doi.org/10.1093/pcp/pcx107
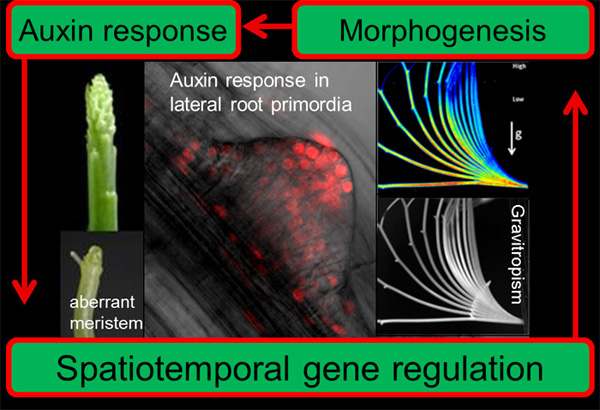
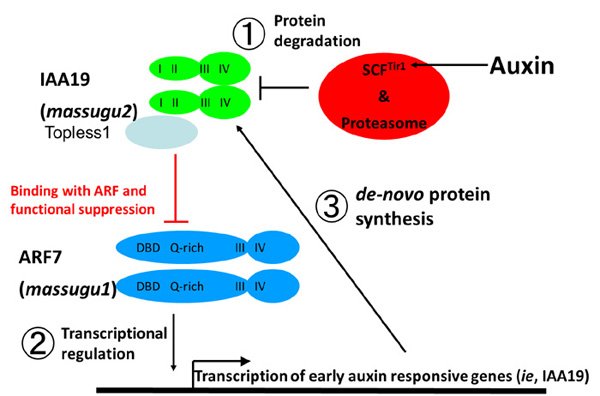
オーキシン非感受性突然変異体,massugu1 (msg1)とmassugu2 (msg2)は共に胚軸の重力応答が弱く,横に寝かせても「まっすぐ」成長します。msg1変異は劣性変異であり,その原因遺伝子はARF7という転写因子をコードしていました。一方,msg2)変異は優性変異であり,原因遺伝子はAUX/IAA19というオーキシンで誘導される遺伝子でした。msg2変異が優性ということは突然変異によって何らかの機能を獲得したかのように考えられます。これはどのようなことなのでしょうか?我々が研究しているAUX/IAA19のオーキシン応答はARF7という転写促進因子の機能阻害が解除されることで起こると考えています(下図中Binding with ARF and functional suppression)。①Tir1/AFBsオーキシンレセプターとAUX/IAA19タンパク質がオーキシン(indole-3-acetic acid)という低分子化合物を介して結合すると,AUX/IAA19タンパク質がユビキチン化され,ユビキチン化されたAUX/IAAタンパク質は速やかに分解されます。②するとAUX/IAA19タンパク質で機能阻害を受けていたARF7転写促進因子が,ターゲット遺伝子群の発現を上昇させることができるようになります。③しかし,ARF7転写促進因子のターゲット遺伝子群の中にAUX/IAA19遺伝子が存在することで,新規のAUX/IAA19タンパク質が作られARF7転写促進因子の機能阻害を起こすようになります。つまり,オーキシン濃度依存的にAUX/IAA19タンパク質の濃度が低下する仕組みでオーキシン濃度依存的に転写調節が行われていると予想しています(下図)。
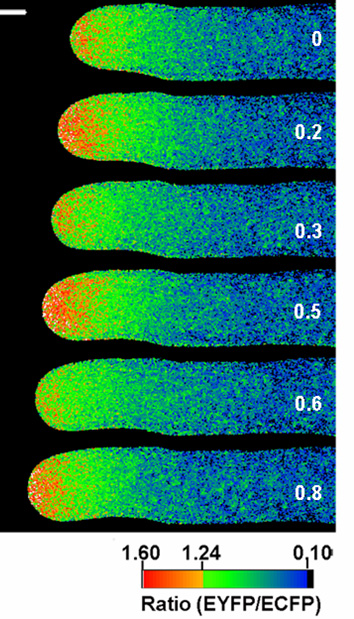
このモデルは時間の進行を考えなければ妥当だと考えられますが,実際オーキシンを投与した時のAUX/IAA19遺伝子の発現は一過的でした。また最近の研究から,植物組織のオーキシン応答は基底状態と定常状態に分けられることがわかってきました。植物の形態形成はいくつもの段階を経ていること,それぞれのステップにオーキシンが関与していることが示されていることから,このような基底状態と定常状態の相互変換が形態形成に重要な役割を演じていると推測されます。私たちはこの基底状態と定常状態の相互変換を「オーキシン応答の再編」と命名し,その分子メカニズムを解明しています。
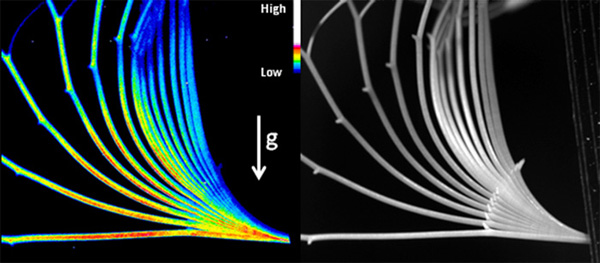
左はシロイヌナズナ花茎を15分間隔で撮影し重ね合わせた写真であり、早期オーキシン応答性遺伝子AUX/IAA19の発現レベルを疑似色で表している。右は同サンプルを赤外線で撮影した明視野像。花茎の重力屈性時、AUX/IAA19は重力方向に偏差的に発現している様子がライブイメージングで明らかになった。